Hai-Rong Xiong,
Yi-Hui Zhang,
Kan Deng,
Qiang Liu,
Yuan-Yuan Liu,
Fan Luo,
We Hou,
Zhan-Qiu Yang ![]()
For correspondence:- Zhan-Qiu Yang Email: zqyang@whu.edu.cn Tel:+862768759136
Received: 9 January 2016 Accepted: 16 June 2016 Published: 31 July 2016
Citation: Xiong H, Zhang Y, Deng K, Liu Q, Liu Y, Luo F, et al. Antiviral activity and mechanism of action of arbidol against Hantaan virus infection. Trop J Pharm Res 2016; 15(7):1439-1445 doi: 10.4314/tjpr.v15i7.12
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the activity and mechanism of action of arbidol against Hantaan virus (HTNV) activity by modulating inflammation via TLR-4 pathway.
Methods: HUVEC cells infected with HTNV 76-118 were treated with serially diluted arbidol solutions at -2h (2 h before viral infection, pre-treatment mode), 0 h (at the same time as viral infection, simultaneous treatment mode) or 2 h (2 h after viral infection, post-treatment mode). The transcript levels of TLR4 were detected by semi-quantitative reverse transcription-PCR (RT-PCR) at 6, 12, 18, and 24 h later. The levels of iNOS and TNF-α were examined using enzyme-linked immunosorbent assay (ELISA).
Results: Pre-treatment with arbidol, rather than simultaneous treatment or post-treatment, effectively inhibited up-regulation of cellular TLR4 ex
Conclusion: These results are the first evidence that arbidol modulates viral PRRs signaling and its consequential inflammatory cytokine/chemokine response during hantavirus infection.
Introduction
Hantaviruses belong to the family, Bunyaviridae, and are associated with two severe forms of hantavirus infections in the human, namely hemorrhagic fever with renal syndrome (HFRS) and hantavirus pulmonary syndrome (HPS), which pose a great threat to human health [1]. Ninety percent of the HFRS cases worldwide were reported in China and mainly caused by two serotypes of hantaviruses-Hantaan virus (HTNV) and Seoul virus (SEOV) [2].
At present, there are no antivirals, or vaccines approved by the Food and Drug Administration (FDA) for the use of hantavirus infection [3]. It is urgent to develop antivirals with novel pharmacological strategies to overcome hantavirus infection. Arbidol is a broad-spectrum antiviral compound that has been shown to have inhibitory effect on influenza virus [4,5], respiratory syncytial virus [6], adenovirus, coxsackievirus [7] and hepatitis C virus infection[8], as well as hantavirus [9]. The antiviral mechanism of arbidol is not completely understood. Early studies indicated that arbidol inhibited membrane fusion of enveloped viruses [10-12]. Arbidol also acts as an immunomodulator, which can induce interferon and activate macrophages in viral infectious patients [13,14]. Our former study indicates that arbidol efficiently inhibits replication of influenza virus and diminishes acute inflammation through modulating the expression of inflammatory cytokines, which indicate the anti-inflammatory activity of arbidol [5].
In a previous study, the present authors demonstrated that arbidol can inhibit HTNV in vitro and in vivo, and it can modulate the level of serum TNF-α in HTNV-infected mouse [9]. Thus, we initiated this study to investigate how arbidol exhibits its anti-HTNV activity by modulating inflammatory effect via TLR-4pathway. Furthermore, we also analyzed this effect in an acute inflammation cell model stimulated by lipopolysaccharide (LPS), a potent inflammatory mediator.
Methods
Cells, virus, and reagents
Arbidol was provided by Shijiazhuang NO.4 Pharmaceutical. Co. Ltd, Hebei, China. HUVEC cells were obtained from CHISCIENTIFIC and were routinely grown in F-12K supplemented with 10 % fetal bovine serum (FBS, GIBCO), 0.1 % L-glutamine, 100 U/mL penicillin and streptomycin. The F-12K containing 2 % FBS was used for maintaining medium after viral infection. The stocks of Hantaan virus, strain 76-118 were obtained from the Institute of Virology, Chinese Academy of Preventive Medicine (Beijing, China) and were propagated on Vero E6 cells. HTNV titration was performed regularly as described previously [9]. Arbidol was provided by Shijiazhuang NO.4 Pharmaceutical. Co. Ltd, Hebei, China. The compound was initially dissolved in 0.05 % dimethyl sulfoxide (DMSO) and then further diluted with maintaining medium.
Experimental design
The cytotoxicity of arbidol on HUVEC cells was determined by quantitative colorimetric MTT assay as described previously [7]. To investigate the dynamic expression pattern of TLR4 after HTNV infection or LPS stimulation, the HUVEC cells were treated with virus or LPS and subsequently incubated for 6, 12, 18, and 24 h.
For arbidol treatment, HUVEC cells were infected with HTNV 76-118 at 100xTCID50 or LPS (100 ng/mL, S1732, Beyotime Institute of Biotechnology, Shanghai, China), and further treated with serially diluted ARB solutions at -2 h (2 h before viral infection, pre-treatment mode), 0 h(at the same time as viral infection, simultaneous treatment mode) or 2 h (2 h after viral infection, post-treatment mode), respectively. After 24 h incubation, the cells were collected for RNA detection by RT-PCR. Four serial dilutions of arbidol (2.5, 5, 10,20μg/mL) were tested in triplicate. Viral control, normal control and solvent control were included in all assays. Cells in extra wells from each group were continuously cultured in corresponding medium for 7 days. Immunofluorescence assay (IFA) was used to detect viral antigen and confirm the successful viral infection in each experiment. In the pre-treatment mode, the supernatant from each wells were collected for iNOS or TNF-α detection.
RT-PCR analysis
Viral RNAs were extracted using TIANamp Virus RNA Kit (Tiangen Biotech Co. Ltd., Beijing) following the manufacturer’s instructions. First-strand cDNA was synthesized by reverse transcription of the total RNA using random primer (Sangon, Shanghai, China) and Moloney murine leukemia virus (M-MLV) Reverse transcriptase(Promega, Beijing, China). Complementary DNA was prepared at 25 oC for 10 min, 42 oC for 60 min, 95 oC for 5 min (Eppendorf, USA).The amplification was performed using the following primer sets: TLR4: 5’-CATTGCTTCTTGCTAAATGCTG-3’ and 5’-GGATTAAAGCTCAGGTCCAGG-3’; GAPDH: 5’-GGTGGTCCTCTGACTTCAACA-3’ and 5’-GTTGCTGTAGCCAAATTCGTTGT-3’, respect-tively. Thirty-two cycles of PCR were carried out in 25 μL reaction mixtures containing 2.5 μL synthesized cDNA. A programmed thermal cycler was set as follows: 94 oC for 180 s, 94 oC for 30s, and 60 oC for 30 s, 72 oC for 15 s, and 72 oC for 5 min. Thereafter, 6 μL of each PCR product was loaded onto an agarose gel containing ethidium bromide and separated by electrophoresis. The gel was scanned using the KODAK Gel Logic 212 PRO Imaging System and analyzed using Bandscan 5.0 software, which calculates the ratio of peak value adsorption of TLR4 to that of GAPDH.
Inducible nitric oxide synthase (iNOS) assay
Inducible nitric oxide synthase (iNOS) activity were determined by a NOS assay kit (Cat no. A014-1, Nanjing Jiancheng Bioengineering Institute, Nanjing, China) following the manufacturer’s instructions. One hundred microliter sample of each group was half diluted and mixed with 100 μL reaction inhibitor solution at room temperature for 3 min. The sample was then incubated with reaction buffer at 37 oC for 15 min. The reaction was terminated by adding the stop buffer and the absorbance was determined at 530 nm. The activity of iNOS was calculated using equation 1:
iNOS activity (U/ml)=(ODsample-ODblank)/(ε × b × 1500) …………………………………….……..(1)
The inhibitory effect of arbidol was evaluated by the following equation 2:
iNOS inhibitory rate=(iNOS unitArbidol-iNOS unitHV)/(iNOS unitNC-iNOS unitHV)×100% .. (2)
TNF-α assay
TNF-α in the supernatant was detected by QuantiKine ELISA kit (R&D Systems, Minneapolis, MN, USA) according to the manufacturer’s instructions. Briefly, 40 μL Assay Diluent and 10 μL sample were successively added in the microplate and incubated at 37 oC for 1 h. Plates were washed with PBS, 200 µL Conjugate and 200 µL substrate solution were added followed by 15 min incubation at 37 oC. The reaction was terminated with stop solution and the absorbance was determined at 450 nm.
Statistical analysis
Data are presented as mean ± standard deviation (SD) and were analyzed by SPSS 17.0 software. One-way ANOVA was used to determine statistical differences for iNOS activity, TNF-α level, and mRNA/β-actin ratio. P < 0.05 was considered to indicate statistical significance.
Results
Cytotoxicity of arbidol on HUVEC cells
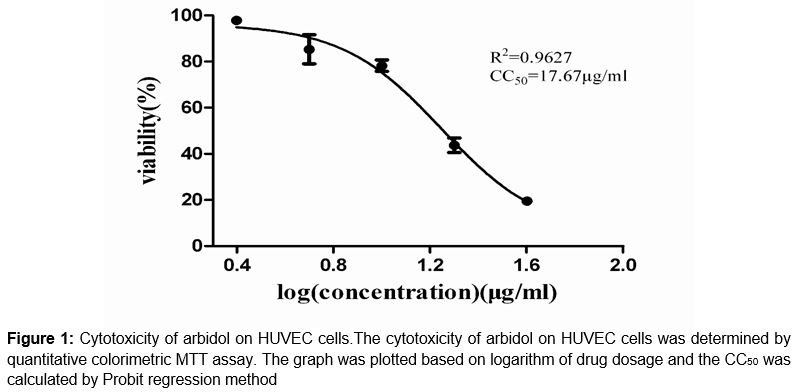
HUVEC cells were treated with arbidol at different concentrations varying from 2.5 to 40 μg/mL and cell viability was investigated by MTT assay (). Subconfluent monolayers treated with arbidol under 10 µg/mL did not show any visible changes in cell morphology or cell density. The CC50 of arbidol was 17.67 μg/mL.
Effect of arbidol on TLR4 expression in HTNV-infected HUVEC cells
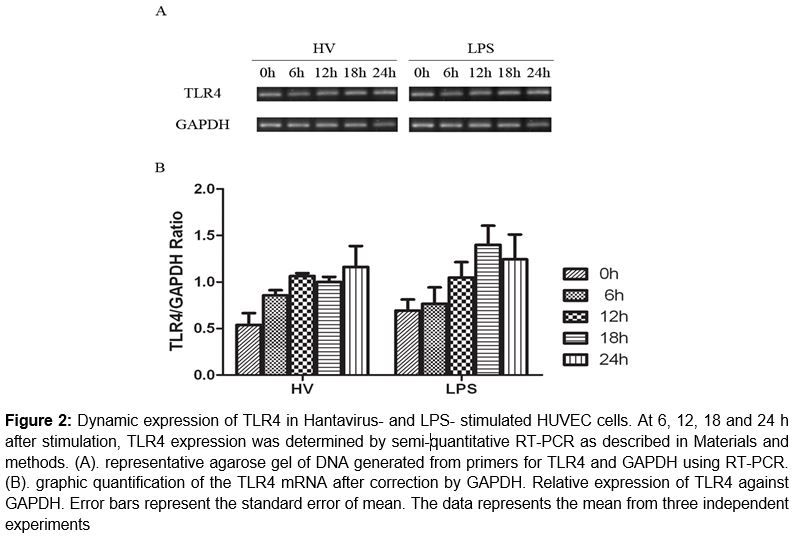
We first analyzed the mRNA expression pattern of TLR4 in HUVEC infected with HTNV and stimulated with LPS. Total RNA was isolated at 6 h, 12 h, 18 h and 24 h post-inoculation and cDNA produced by reverse transcription were subjected to PCR analysis. As shown in , TLR4 was expressed at detectable level in both HTNV- and LPS- stimulated cells and the expression level after HTNV- and LPS-stimulation are significantly different from the initial normal expression (p < 0.05, except of 6 h in LPS group). The expression pattern in these two groups were similar (p = 0.2815).
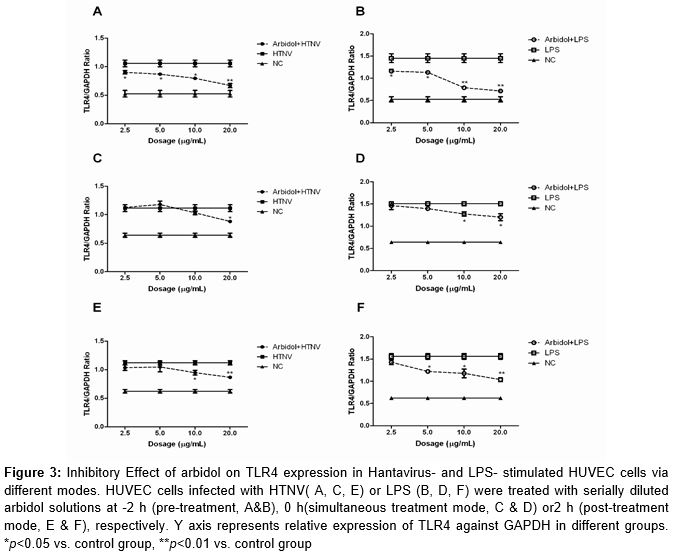
We further investigated the effect of arbidol on TLR4 expression in HTNV-infected and LPS-stimulated HUVEC cells. Arbidol treatment was applied to different modes as described in Material and Methods. In the pre-treatment mode, arbidol significantly inhibited the up-regulated TLR4 expression induced by hantavirus infection or LPS stimulation (A & B). This effect increased with increase in drug concentration. The 20 μg/mL arbidol inhibited around 40 ± 6.1 % TLR4 expression in HTNV-infected cells, and 50 ± 1.8 % in the LPS group (p < 0.05). In the simultaneous treatment mode, the inhibitory effect was observed only at 20 μg/mL in hantavirus infection group (p = 0.041, C) or 10.0 μg/mL and 20 μg/mL in LPS stimulation group (p = 0.025, p = 0.028, D), and not than in the low-concentration groups. In the post-treatment mode, TLR4 expression in HUVEC cells treated with arbidol at 20 μg/mL showed a significant reduction (p < 0.01, E & F).
Taken together, pre-treatment with arbidol inhibited the increased TLR4 expression due to hantavirus infection or LPS stimulation in a concentration-dependent manner. High dosage of arbidol also showed inhibitory effects in the other treatment modes.
Effect of arbidol on iNOS in HTNV-infected HUVEC cells
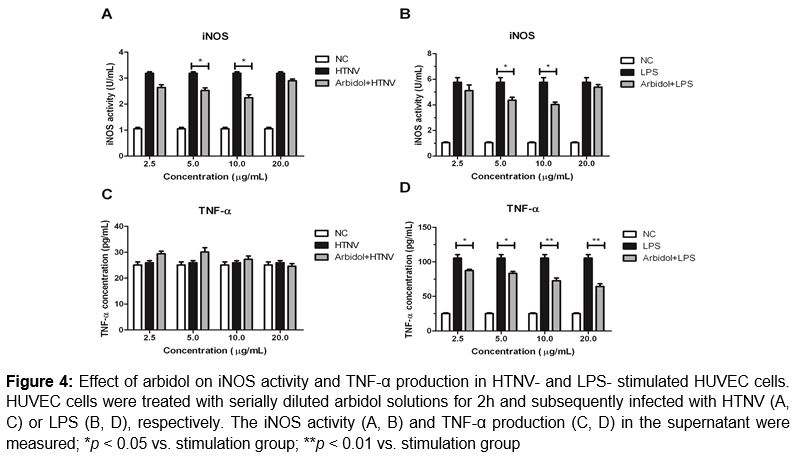
To test whether arbidol has an impact on NO production, HUVEC cells were first pre-treated with different concentrations of arbidol for 2 h, and then infected with HTNV or stimulated with LPS. Twenty four hours later, the amount of nitrite in the supernatant was measured. As shown in A & B, elevated levels of nitrite were detected in the supernatant from HUVEC cells after HTNV or LPS stimulation, with approximately 2.8- or 5-fold increase compared to non-infected cells (p < 0.01). Arbidol treatments at concentration of 5 and 10 μg/mL inhibited the elevated iNOS with approximately 30.9 ± 8.1 and 44.1 ± 9.4 % respectively (p < 0.05).There were no inhibitory effect observed in the high concentration group (20 μg/mL) or low concentration group (2.5 μg/mL).
Effect of arbidol on TNF-α in HTNV-infected HUVEC cells
It is known that TNF-α induces nitric oxide (NO) through the induction of NOS, thus we tested whether TNF-α was involved in the anti-viral or anti-inflammatory effect of arbidol in HTNV-infected or LPS-stimulated HUVEC cells. The results showed that LPS stimulation, rather than HTNV infection, could induce TNF-α production 3.7 times higher than normal control group (p < 0.01, C & D). Furthermore, arbidol could decrease the elevated TNF-α levels induced by LPS stimulation in a concentration-dependent manner (D).
Discussion
Arbidol is a small antiviral compound with a broad-spectrum inhibitory effect. We have previously shown that arbidol could inhibit HTNV in vitro and in vivo. Here, our results demonstrated that arbidol could inhibit the up-regulation of cellular TLR4 expression as well as iNOS induced by HTNV infection, and also inflammation stimulated with LPS.
It has been reported that HTNV induces TLR4 expression in virus-infected EVC-304 cells, leading to enhanced production of IFN-β, IL-6 and TNF-α [15]. Arbidol possesses comprehensive pharmacological properties and is also known as an immunomodulant and interferon inducer. However, Boriskin et al found that arbidol inhibited acute HCV infection without affecting RIG-I and IFN signaling pathway [8], which implied other antiviral mechanisms of arbidol besides interferon induction. We found that arbidol inhibited the increased TLR4 expression due to hantavirus infection or LPS stimulation, especially in the pre-treatment group in a concentration-dependent manner. This result suggests that arbidol may regulate the TLR4-mediated innate immune response in the HTNV-infected HUVECs.
We further tested if arbidol could modulate the inflammatory cytokines/chemokines produced by antiviral innate response during HTNV infection. Increased amounts of iNOS have been detected in patients with hantavirus-related disease, which could facilitate immunopathological processes during the infection [16,17]. Administration of a specific iNOS inhibitor significantly improved survival in hantavirus-infected murine model with HCPS-like disease [16]. We found that median concentration of arbidol could attenuate the elevated iNOS level in HTNV- or LPS- stimulated HUVECs, which suggests that arbidol may alleviate the pathological consequences of HTNV infection/acute inflammation via inhibition of iNOS generation.
It has been reported that hantavirus infection can up-regulate the production of TNF-α in HFRS patients [18,19]. Niikura et al demonstrated that TNF-α could extend the duration of increased vascular permeability of the HTNV-infected HUVEC [20]. It could also induce nitric oxide (NO) through induction of NOS [21]. We tried to examine if arbidol could inhibit the production of iNOS via suppression of TNF-α expression. However, our results showed that arbidol could decrease the elevated TNF-α levels induced by LPS stimulation in a concentration-dependent manner. We did not detect any change of TNF-α in the supernatant of HTNV-infected HUVECs, nonetheless, other in vitro studies reported that infection of immature dendritic cells or human alveolar macrophages with hantavirus weakly up-regulated TNF-α production.
We assume that the HTNV infection of HUVEC itself did not induce TNF-α production as it did in other cell lines or the production level is too low and beyond the detection range in our system. Previously we have found that arbidol could modulate TNF-α response in HTNV-infected suckling mice [9], we presumed that arbidol may modulate the inflammatory chemokine in an integral animal infection model rather than cell infection model.
Conclusion
Arbidol inhibits TLR4 expression and iNOS production induced by HTNV infection in HUVECs, which represents one of the virus-sensing pattern-recognition receptors (PRRs) and its consequential inflammatory cytokines/chemokines. Further experiments are needed to investigate the in-depth mechanism of action of aribidol.
Declarations
Acknowledgement
References
Archives


News Updates